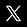|
Enzyme
Un article de Wikipédia, l'encyclopédie libre.
Aller à : Navigation, rechercher
Figure 1: Diagramme d'une réaction catalytique qui montre l'énergie (E) requise à différentes étapes suivant l'axe du temps (t). Les substrats (A et B) en conditions normales requièrent une quantité d'énergie considérable (E1) pour atteindre l'état de transition A...B, à la suite duquel le produit de réaction (AB) peut se former. L'enzyme (E) crée un microenvironnement dans lequel A et B peuvent atteindre l'état de transition (A...E...B) plus facilement, réduisant ainsi la quantité d'énergie requise (E2). Comme il est plus aisé d'atteindre un niveau énergétique inférieur, la réaction peut avoir lieu plus fréquemment, ce qui se traduit par une vitesse de réaction accrue.
Une enzyme (parfois utilisé au masculin) est une molécule (protéine ou ARN dans le cas des ribozymes) permettant d'abaisser l'énergie d'activation d'une réaction et d'accélérer jusqu'à des millions de fois les réactions chimiques du métabolisme se déroulant dans le milieu cellulaire ou extracellulaire sans modifier l'équilibre formé. Les enzymes agissent à faible concentration et elles se retrouvent intactes en fin de réaction : ce sont des catalyseurs biologiques (ou biocatalyseurs). La première enzyme fut découverte par Anselme Payen et Jean-François Persoz en 1833[1]. Après avoir traité un extrait aqueux de malt à l'éthanol, ils ont précipité une substance sensible à la chaleur. Cette substance était capable d'hydrolyser l'amidon : ils l'ont nommée diastase (étym. : diastasis = séparation) car elle séparait le sucre soluble de l'amidon. On l'appelle aussi amylase α (amylase alpha).
La nomenclature des enzymes n'est pas standardisée mais, par convention, elle se compose le plus souvent d'un radical proche du substrat ou du produit de la catalyse suivi du suffixe « ase ». Par exemple :
La glucose-oxydase est une enzyme qui catalyse l'oxydation du glucose.
L'amidon-synthétase catalyse la synthèse de l'amidon.
Une enzyme, comme toute protéine, est synthétisée par les cellules vivantes à partir des informations codées dans l'ADN ou dans l'ARN dans le cas de certains virus. Il existe plus de 3 500 enzymes différentes répertoriées.
Les premières enzymes isolées furent d'abord nommées ferments solubles, diastases ou zymases.
Il existe deux grandes catégories d'enzymes :
les enzymes purement protéiques (qui ne sont constituées que d'acides aminés)
les enzymes en deux parties, une partie protéique (« l'apoenzyme ») et une partie non protéique (« le cofacteur »), appelées « holoenzymes »[2]. Dans ce cas, sans la coenzyme, la catalyse de la réaction ne peut avoir lieu.
Exemple : L'aminoacyl-ARNt synthétase (enzyme fixant l'acide aminé sur l'ARN transfert) a besoin d'ATP (coenzyme) pour fixer l'acide aminé sur l'ARNt.
Sommaire [masquer]
1 Site actif
2 « Vitesse initiale » de réaction enzymatique
3 Régulation de l'activité enzymatique
4 Classement et dénomination des enzymes
5 Terminologie
6 Pathologie
7 Notes et références
8 Voir aussi
8.1 Articles connexes
8.2 Liens externes
DOCUMENT WIKIPEDIA LIEN |