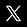|
|
|
|
 |
|
NEURONE |
|
|
| |
|
| |
NEURONE
Cellule de base du tissu nerveux, capable de recevoir, d'analyser et de produire des informations. (La partie principale, ou corps cellulaire du neurone, est munie de prolongements, les dendrites et l'axone.)
Le neurone, ou cellule nerveuse, est l'unité fonctionnelle du système nerveux. Sa configuration cellulaire spécifique le rend capable de générer, de transmettre et/ou de recevoir des informations sous forme de signaux électriques (influx nerveux).
Le système nerveux est très complexe : ses quelque cent milliards de cellules (1011, à un facteur de 10 près) peuvent être classées en plus de 1 000 catégories, chacune comprenant plusieurs sous-classes selon des critères incluant leur taille, leurs arborisations, les contacts afférents ou efférents. Cette apparente complexité masque en fait une grande similitude fonctionnelle, la particularité de chaque cellule reposant principalement sur sa position dans un circuit donné.
On peut dès lors essayer de mieux comprendre le système nerveux à partir de propriétés simples des cellules qui le composent : les mécanismes qui donnent naissance aux potentiels d'action neuronaux, les modes de transmission synaptique, les interactions fondamentales entre neurones et cellules gliales.
En effet, les neurones sont entourés d'autres cellules : les cellules gliales (du grec gloios, « glu ») – elles ont longtemps été considérées comme de simples éléments de remplissage entre les neurones. On compte environ dix fois plus de cellules gliales que de neurones, et il est clair aujourd'hui que leurs fonctions, essentielles à l'activité neuronale, sont multiples.
Les compartiments du neurone
Le neurone est constitué de quatre grandes régions : le corps cellulaire, ou soma, les dendrites, les axones et les terminaisons présynaptiques. Chacune de ces différentes régions joue un rôle bien défini dans l'initiation et le transport de l'information.
Le soma, qui contient le noyau du neurone, en est le centre métabolique, car il comprend aussi toute la machinerie de synthèse de ses différents constituants. Du corps cellulaire partent deux sortes de prolongements, les dendrites et l'axone.
Les dendrites, qui se ramifient au point de former un arbre touffu autour du corps cellulaire (arbre dendritique), sont les voies par lesquelles l'information arrive. Elles sont le siège d'une activité métabolique intense et d'une synthèse protéique active : la microscopie électronique permet d'y distinguer des mitochondries en abondance, ainsi qu'un réticulum endoplasmique rugueux, porteur de nombreux ribosomes.
L'axone, prolongement au diamètre constant (de 0,2 à 20 μm), peut atteindre 1 m de longueur. Il est la voie privilégiée de sortie de l'information, et, contrairement aux dendrites, l'activité métabolique y est peu importante. L'axone transporte en revanche les macromolécules stockées dans des vésicules, ou organelles, au sein du corps cellulaire. Dans certains cas, ce transport permet une maturation de la molécule, importante pour sa fonction.
Près de sa terminaison, l'axone se divise en fines ramifications, les terminaisons présynaptiques. Ces dernières sont le site de stockage des neurotransmetteurs, qui vont permettre le transfert de l'information aux dendrites du neurone postsynaptique.
L’influx nerveux
L'une des propriétés essentielles du neurone est sa capacité à produire, puis à acheminer loin du corps cellulaire, une information sous la forme d'un groupe d'impulsions électriques, les potentiels d'action.
Décrite dès 1849 par le biologiste allemand Emil Du Bois-Reymond (1818-1896), cette aptitude résulte des propriétés de la membrane cellulaire du neurone et des protéines qu'elle contient. Les protéines membranaires des cellules de l'organisme peuvent être regroupées en cinq grandes familles : les pompes, les canaux, les récepteurs, les enzymes et les protéines de structure.
Les échanges d'ions entre le neurone et son milieu
Les pompes utilisent l'énergie produite à partir de la dégradation des sucres pour déplacer activement des ions et d'autres molécules contre leur gradient de concentration (un gradient est créé de fait par les différences de concentration d'une substance de part et d'autre d'une membrane ; celle-ci peut être traversée passivement – sans nécessiter de pompes – par les ions, du milieu le plus concentré vers le moins concentré, c'est-à-dire dans le sens du gradient). La composition ionique du milieu intracellulaire est différente de celle du milieu extracellulaire, et ce pour toutes les cellules de l'organisme. À l'intérieur d'un neurone, il y a dix fois plus de potassium et dix fois moins de sodium qu'à l'extérieur.
La pompe Na-K-ATPase échange trois ions sodium de l'intérieur contre deux ions potassium de l'extérieur. Ces échanges ioniques induisent une différence de potentiel au niveau de la membrane ; celle-ci a un potentiel d'environ 60 mV.
Le potentiel transmembranaire
Comme le milieu intérieur, concentré en protéines chargées négativement, est négatif, et que le milieu extracellulaire, choisi comme référence, est à zéro, le potentiel de repos d'un neurone se situe à − 60 mV. Cette valeur est prise comme base à partir de laquelle les variations traduisent l'apparition d'une information.
Toute augmentation (en valeur absolue) du potentiel transmembranaire (de − 60 à − 70 mV, par exemple) est une hyperpolarisation ; inversement, une diminution de potentiel (de − 60 à − 50 mV, par exemple) est une dépolarisation.
L'hyperpolarisation éloigne du seuil d'apparition d'un potentiel d'action, tandis que la dépolarisation est l'étape initiale pouvant donner naissance, si elle est suffisamment intense, à la « décharge » du neurone : le potentiel d'action. Lorsqu'un signal atteint le neurone, il en résulte une hyper- ou une dépolarisation.
Dans le premier cas, on parle de signal inhibiteur, tandis qu'il est excitateur dans le second. Le stimulus peut être de toute nature : lumière, bruit, odeur, étirement musculaire, molécule chimique libérée par un autre neurone, etc. Il en résulte une perturbation du potentiel de repos de faible amplitude (moins de 10 mV), locale et graduée : locale, car la résistance passive de la membrane limite la diffusion de la perturbation ; graduée, car le changement de potentiel est proportionnel à l'intensité de la stimulation ; on parle de potentiel de récepteur et/ou de potentiel synaptique.
L'ensemble des potentiels qui atteignent un même neurone est intégré au niveau d'une zone spécialisée de la membrane, appelée trigger zone, ou zone gâchette. C'est là que la sommation des hyper- et/ou des dépolarisations élémentaires se transforme ou non en un potentiel d'action.
La naissance du potentiel d'action
Le potentiel d'action est une dépolarisation ample (jusqu'à 110 mV), brève (1/1 000 s), générée selon la loi du « tout ou rien », et propagée activement le long du neurone et de l'axone sans diminution d'amplitude.
Dans le courant des années 1950, Alan L. Hodgkin, Andrew F. Huxley et Bernard Katz démontrèrent, sur l'axone géant de calmar, que la propagation de l'influx nerveux coïncidait avec un brusque changement de la perméabilité membranaire aux ions sodium (Na+) et potassium (K+). Au repos, la membrane est principalement perméable au potassium (on parle de conductance potassique), qui passe par des canaux dits « de fuite ». Hodgkin et Katz avancèrent l'hypothèse que l'influx nerveux modifie la conductance, car il entraîne l'ouverture brutale des canaux sodiques sensibles au potentiel de la membrane. Ces derniers s'ouvrent dès que la différence de voltage atteint un seuil de − 55 mV, et laissent entrer massivement le sodium dans le sens de son gradient ; cette entrée est à l'origine du potentiel d'action, le flux de sodium ouvre davantage de canaux Na+, facilitant ainsi le passage d'autres ions sodium.
La dépolarisation s'amplifie alors jusqu'à activer les canaux K+, sensibles à des valeurs de potentiel proches de 0 mV : le flux sortant de potassium compense le flux entrant de sodium. Le potentiel membranaire reprend sa valeur initiale. Les pompes Na-K-ATPase rétablissent les gradients initiaux au cours d'une phase dite « réfractaire », durant laquelle aucun potentiel d'action ne peut être produit.
La propagation de l'influx nerveux
À partir de la trigger zone, le potentiel d'action avance vers l'extrémité de l'axone à grande vitesse. Toutefois, en raison des pertes dues aux résistances de membrane, ce potentiel doit être régénéré de façon active tout au long de son parcours. Au cours de l'évolution, deux stratégies ont été mises en place par les organismes vivants pour augmenter la vitesse de conduction de l'information le long de l'axone.
Chez certaines espèces, l'augmentation du diamètre de la fibre a été poussée à l'extrême : il atteint 1 mm chez le calmar géant, et peut être vu à l'œil nu – ce caractère évolutif, à l'origine d'un encombrement spatial important, n'est pas compatible avec le nombre de neurones observés dans le cerveau des vertébrés supérieurs.
Chez les mammifères, la vitesse de propagation de l'influx nerveux est augmentée par la présence d'une gaine de myéline autour de l'axone ; cette enveloppe protéique et lipidique, qui s'enroule sur une dizaine de couches, est synthétisée par les cellules gliales spécialisées du système nerveux central, les oligodendrocytes, et par celles du système nerveux périphérique, les cellules de Schwann. La diminution de la résistance passive de la membrane est telle que le potentiel d'action est transporté cent fois plus rapidement : il avance alors de façon saltatoire, en bondissant le long de l'axone, d'un nœud de Ranvier (zone non myélinisée à la jonction entre deux cellules myélinisantes) à l'autre.
Le transfert de l’information d’une cellule à l’autre
Le transfert de l’influx nerveux d’un neurone à l’autre, ou, en fin de circuit, d’un neurone à une cellule effectrice (cellule musculaire par exemple), se fait au niveau d’une zone de jonction appelé synapse. Il existe deux types de synapses : les synapses chimiques et les synapses électriques. Les premières, majoritaires, font appel à des molécules messagers appelées neuromédiateurs.
→ synapse
Les cellules gliales, partenaires du neurone
Les oligodendrocytes et les cellules de Schwann, nécessaires à la transmission rapide du potentiel d'action, forment une gaine isolante, la myéline ; sa destruction par la maladie (sclérose en plaques) induit une grave perturbation de l'activité neuronale, qui peut conduire à la disparition du neurone.
Les cellules de la microglie, qui assurent la défense immunitaire du système nerveux, sont des cibles privilégiées pour les virus, comme celui responsable du sida. De plus, elles ont un rôle important lors du développement embryonnaire, en éliminant les cellules et les terminaisons surnuméraires, et lors des phénomènes de sénescence.
Les cellules endothéliales, qui bordent les vaisseaux cérébraux, sont responsables de la barrière hémato-encéphalique ; celle-ci isole le système nerveux de la circulation sanguine générale, le protégeant ainsi de l'arrivée de nombreux toxiques et agents infectieux.
Les astrocytes représentent la principale population gliale, et leurs rôles apparaissent aujourd'hui multiples. Lors du développement, ils participent au guidage et au positionnement des neurones. Ils possèdent une capacité de captation rapide du potassium extracellulaire, permettant une rapide repolarisation neuronale et la régénération du potentiel d'action. Ils approvisionnent en substrats énergétiques le neurone, isolé de la circulation sanguine. De plus, ils expriment de nombreux récepteurs pour les neurotransmetteurs libérés par les neurones, les rendant ainsi aptes à prendre une place – qui reste à mieux comprendre – dans les processus de transmission de l'information au sein du système nerveux.
Neurone et intelligence artificielle
Imiter le système nerveux au moyen d'une machine n'est pas une idée nouvelle, mais ce projet connaît aujourd'hui un fort développement grâce à l'informatique, l'ordinateur permettant de tester, de simuler les hypothèses émises par les chercheurs. Et c'est à l'échelle de la cellule nerveuse, ou neurone, que les chercheurs en neuro-mimétisme ont décidé de se placer pour réaliser la copie de la structure du système nerveux. L'ordinateur sert en particulier à faire fonctionner les modèles construits par les mathématiciens à partir des observations et des réflexions fournies par les physiologistes. Ce modèle mathématique du neurone, que l'on appelle le « neurone formel », est dupliqué pour constituer un réseau.
Pour en savoir plus, voir l'article neurone formel.
DOCUMENT larousse.fr LIEN |
| |
|
| |
|
|
|
Les préférences auditives des nouveau-nés sont influencées par lâexpérience émotionnelle associée aux sons perçus par leur mère au cours de la gestati |
|
|
| |
|
| |
Les préférences auditives des nouveau-nés sont influencées par l’expérience émotionnelle associée aux sons perçus par leur mère au cours de la gestation
L’expérience sensorielle prénatale, en particulier les expériences auditives, sont une source de mémoires pour les fœtus. Ainsi, les nouveau-nés de différentes espèces, y compris l’Homme, distinguent la voix de leur mère de celle d’autres individus. Des chercheurs des laboratoires Physiologie, environnement et génétique pour l'animal et les systèmes d'élevage et Ethologie animale et humaine, apportent pour la première fois une preuve expérimentale chez des porcelets que la contiguïté temporelle entre des stimuli sensoriels et des réactions émotionnelles maternelles peut être une source d’apprentissage associatif pour le fœtus. Cette étude a été publiée le 18 novembre dans la revue Scientific Reports.
Les chercheurs ont testé si l’association de sons (voix humaine lisant un texte) avec une expérience émotionnelle de valence positive ou négative chez des truies gestantes avait un impact sur la réaction des porcelets à ces mêmes sons juste après la naissance et dans les semaines suivantes.
Trente truies et leurs portées ont été étudiées : les truies recevaient quotidiennement et alternativement un traitement positif et un traitement négatif pendant leur dernier mois de gestation, chacun étant associé avec une voix humaine particulière. Après la naissance des porcelets, ceux-ci ont été soumis à des tests de séparation auxquels étaient associés la diffusion d’une ou des deux voix. Les résultats montrent bien que cette expérience de la mère gestante a une influence sur les réactions postnatales des porcelets à ces sons : 1) dès le premier test à l’âge de deux jours (mais également encore à l'âge de trois semaines), les porcelets pour qui ces voix sont “familières” sont moins stressés par la séparation que les porcelets témoins qui n'ont pas été exposés à ces voix avant leur naissance : il y a donc bien eu audition et mémorisation in utero ; 2) cet effet est généralisé à toute voix humaine puisqu’ils sont aussi davantage “rassurés” en entendant une voix étrangère ; 3) et surtout, ils sont plus stressés en entendant la voix humaine associée par la mère à une émotion négative que lorsqu'ils sont isolés avec la voix humaine associée par la mère à une émotion positive ou en absence de voix. Cette étude apporte la première démonstration d’un apprentissage in utero d’une association entre un stimulus sensoriel et l’état émotionnel de la mère. Ces résultats ouvrent tout un pan de recherche qui va bien au-delà de l’espèce porcine car ils invitent à reconsidérer l’impact des expériences prénatales à travers le filtre des ressentis maternels.
En savoir plus
* Postnatal auditory preferences in piglets differ according to maternal emotional experience with the same sounds during gestation.
Tallet C, Rakotomahandry M, Guérin C, Lemasson A, Hausberger M.
Sci Rep. 2016 Nov 18;6:37238. doi: 10.1038/srep37238.
Contact chercheur
* Céline Tallet
Physiologie, Environnement et Génétique
pour l'Animal et les Systèmes d'Élevage
INRA UMR 1348- Université de Rennes 1
INRA St Gilles Domaine de la Prise
35590 SAINT GILLES Tel: 02 23 48 50 53
* Martine Hausberger
Ethologie animale et Humaine (Ethos)
CNRS UMR 6552 - Université de Rennes 1
Campus de Beaulieu
263 Avenue du Général Leclerc
35042 Rennes cedex
Tel: 02 23 23 48 28
* Alban Lemasson
Ethologie animale et Humaine (Ethos)
CNRS UMR 6552 - Université de Rennes 1
Campus de Beaulieu
263 Avenue du Général Leclerc
35042 Rennes cedex
Tel: 02 23 23 68 20
DOCUMENT cnrs LIEN
|
| |
|
| |
|
|
|
Impact de lâapprentissage de la lecture sur le cerveau |
|
|
| |
|
| |
Impact de l’apprentissage de la lecture sur le cerveau
COMMUNIQUÉ | 09 NOV. 2010 - 17H23 | PAR INSERM (SALLE DE PRESSE)
Pour la première fois, des images détaillées de l’impact de l’apprentissage de la lecture sur le cerveau ont été obtenues par une équipe internationale de chercheurs. En comparant l’activité cérébrale d’adultes analphabètes avec celle de personnes alphabétisées durant l’enfance ou à l’âge adulte ces chercheurs ont démontré l’emprise massive de la lecture sur les aires visuelles du cerveau ainsi que sur celles utilisées pour le langage parlé. Coordonnée par Stanislas Dehaene (Collège de France, Unité CEAInserm- Université Paris Sud 11 de Neuroimagerie Cognitive, NeuroSpin/I²BM) et Laurent Cohen (Inserm, AP-HP, Université Pierre et Marie Curie), cette étude a impliqué des équipes brésiliennes, portugaises, et belges. Ces résultats sont publiés en ligne le 11 novembre par la revue Science.
L’acquisition de la lecture soulève plusieurs questions scientifiques importantes quant à son influence sur le fonctionnement cérébral. L’écriture est une invention trop récente pour avoir influencé l’évolution génétique humaine. Son apprentissage ne peut donc reposer que sur un « recyclage » de régions cérébrales préexistantes, initialement dédiées à d’autres fonctions mais suffisamment plastiques pour se réorienter vers l’identification des signes écrits et leur mise en liaison avec le langage parlé (1). C’est dans ce cadre que les chercheurs essaient de mieux comprendre l’impact de l’apprentissage de la lecture sur le cerveau.
Pour cela, ils ont mesuré, par IRM fonctionnelle (2), l’activité cérébrale d’adultes volontaires diversement alphabétisés, dans l’ensemble du cortex, avec une résolution de quelques millimètres, tandis qu’ils leur présentaient toute une batterie de stimuli : phrases parlées et écrites, mots et pseudo-mots parlés, visages, maisons, objets, damiers…63 adultes ont participé à l’étude : 10 personnes analphabètes, 22 personnes non-scolarisées dans l’enfance mais alphabétisées à l’âge adulte, et 31 personnes scolarisées depuis l’enfance. La recherche a été menée en parallèle au Portugal et au Brésil, pays dans lesquels, voici quelques dizaines d’années, il était encore relativement fréquent que des enfants ne puissent pas aller à l’école uniquement en raison de leur environnement social (isolement relatif, milieu rural). Tous les volontaires étaient bien intégrés socialement, en bonne santé, et la plupart avaient un emploi. Les études ont été réalisées avec des imageurs IRM à 3 Tesla au centre NeuroSpin (CEA Saclay) pour les volontaires portugais et au centre de recherches en neurosciences de l’hôpital Sarah Lago Norte à Brasilia3 pour les volontaires brésiliens. Grâce à ces travaux les chercheurs apportent des éléments de réponse à plusieurs questions essentielles.
Comment les aires cérébrales impliquées dans la lecture se transforment-elles sous l’influence de l’éducation ?
En comparant directement l’évolution de l’activation cérébrale en fonction du score de lecture (nul chez les analphabètes et variable dans les autres groupes), les chercheurs ont montré que l’impact de l’alphabétisation est bien plus étendu que les études précédentes ne le laissaient penser.
* Apprendre à lire augmente les réponses des aires visuelles du cortex, non seulement dans une région spécialisée pour la forme écrite des lettres (précédemment identifiée comme la « boîte aux lettres du cerveau »), mais aussi dans l’aire visuelle primaire.
*
* La lecture augmente également les réponses au langage parlé dans le cortex auditif, dans une région impliquée dans le codage des phonèmes (les plus petits éléments significatifs du langage parlé, comme « b » ou « ch »). Ce résultat pourrait correspondre au fait que les analphabètes ne parviennent pas à réaliser des jeux de langage tels que la délétion du premier son d’un mot (Paris→aris).
*
* La lecture induit également une extension des aires du langage et une communication bidirectionnelle entre les réseaux du langage parlé et écrit : chez un bon lecteur, voir une phrase écrite active l’ensemble des aires du langage parlé, entendre un mot parlé permet de réactiver rapidement son code orthographique dans les aires visuelles. Chez les personnes qui n’ont pas appris à lire, le traitement du langage est moins flexible et strictement limité à la modalité auditive.
*
*
À quoi servent les aires cérébrales impliquées dans la lecture avant qu’une personne n’apprenne à lire ? L’apprentissage de la lecture implique-t-il toujours un gain de fonction, ou bien l’augmentation des réponses aux mots s’accompagne-t-elle de diminutions des réponses à d’autres catégories de connaissances ?
Chez les analphabètes l’aire visuelle de l’hémisphère gauche qui, chez les lecteurs, décode les mots écrits répond à une fonction proche : la reconnaissance visuelle des objets et des visages. Dans cette région, au cours de l’apprentissage, la réponse aux visages diminue légèrement à mesure que la compétence de lecture augmente, et l’activation aux visages se déplace partiellement dans l’hémisphère droit. Le cortex visuel se réorganise donc, en partie, par compétition entre l’activité nouvelle de lecture et les activités plus anciennes de reconnaissance des visages et des objets. Aujourd’hui, on ne sait pas si cette compétition entraîne des conséquences fonctionnelles pour la reconnaissance ou la mémoire des visages.
Les modifications cérébrales liées à l’alphabétisation peuvent-elles se produire à l’âge adulte ? Ou bien existe-t-il une « période critique » pour cet apprentissage dans la petite enfance ?
La très grande majorité des effets de l’apprentissage de la lecture sur le cortex sont visibles autant chez les personnes scolarisées dans l’enfance que chez celles qui ont suivi des cours d’alphabétisation à l’âge adulte. Bien entendu, ces dernières n’atteignent que rarement les mêmes performances de lecture, mais cette différence pourrait n’être due qu’à leur moindre entraînement. À performances de lecture égales, il n’existe pratiquement pas de différences mesurables entre les activations cérébrales des personnes qui ont appris à lire dans l’enfance ou à l’âge adulte. Les circuits de la lecture restent donc plastiques tout au long de la vie.
Ces résultats soulignent l’impact massif de l’éducation sur le cerveau humain. Ils nous rappellent également que l’immense majorité des expériences d’IRM cérébrale portent sur le cerveau éduqué et que l’organisation cérébrale en l’absence d’éducation constitue un immense territoire largement inexploré.
Un aperçu des vastes réseaux cérébraux dont l’activité augmente avec le score de lecture, en réponse à des phrases écrites. Dès qu’une personne sait lire, la réponse aux mots écrits augmente rapidement dans diverses aires visuelles, dont l’une est spécialisée dans l’analyse de la forme des lettres (graphe de droite). De plus, l’ensemble des régions de l’hémisphère gauche impliquées dans le traitement du langage parlé (médaillon) devient susceptible de s’activer également en réponse au langage écrit.
DOCUMENT inserm LIEN |
| |
|
| |
|
|
|
Une enzyme cruciale enfin démasquée |
|
|
| |
|
| |
Une enzyme cruciale enfin démasquée
| 20 NOV. 2017 - 15H19 | PAR INSERM (SALLE DE PRESSE)
BIOLOGIE CELLULAIRE, DÉVELOPPEMENT ET ÉVOLUTION | CANCER | NEUROSCIENCES, SCIENCES COGNITIVES, NEUROLOGIE, PSYCHIATRIE
Après 40 ans de recherche, des chercheurs du CEA, du CNRS, de l’Université Grenoble Alpes, de l’Université de Montpellier et de l’Inserm ont enfin démasqué l’enzyme responsable de la détyrosination de la tubuline. Surprise : ce n’est pas une enzyme mais deux qui ont été découvertes capables de modifier ce composant essentiel du squelette de la cellule. Ces travaux ouvrent de nouvelles pistes pour mieux comprendre le rôle de la tubuline dont les altérations accompagnent cancers, maladies cardiaques et défauts neuronaux. Ces résultats sont publiés le 16 novembre 2017 dans la revue Science.
Une collaboration internationale impliquant des chercheurs du CEA, du CNRS, de l’Inserm, de l’Université Grenoble Alpes, de l’Université de Montpellier et de l’Université de Stanford[1] a identifié une enzyme, la Tubuline CarboxyPeptidase (TCP), qui est responsable d’une transformation biochimique des microtubules cellulaires, la détyrosination. La détyrosination est une réaction biologique consistant à supprimer l’acide aminé terminal tyrosine[2], de la tubuline α, un composant des microtubules. Alors qu’elle était recherchée depuis quatre décennies, les biologistes ont réussi à isoler cette protéine par purification et ont ensuite apporté les preuves de son activité cellulaire.
Les microtubules contribuent à des fonctions cellulaires essentielles
Les microtubules sont des fibres dynamiques présentes dans toutes les cellules. Formés par l’assemblage de deux protéines (tubuline α et tubuline β), les microtubules assurent de nombreuses fonctions. Ils séparent les chromosomes destinés aux deux cellules filles lors de la division cellulaire, ils contribuent à la polarité des cellules, à la morphologie et à la migration cellulaire. Ils forment des sortes de rails sur lesquels sont transportés des constituants cellulaires tels que des protéines ou des brins d’ARN.
Ces fonctions cellulaires sont régulées grâce à l’existence de « signaux » présents à la surface des microtubules. Ces signaux sont des modifications biochimiques des acides aminés (appelées modifications post-traductionnelles car elles ont lieu après la synthèse de la protéine) qui sont réalisées par plusieurs enzymes qui, ici, modifient les tubulines.
L’enzyme TCP, identifiée après 40 ans de mystère
L’activité de l’une de ces enzymes a été mise en évidence pour la première fois en 1977 par des chercheurs argentins qui lui donnent le nom de TCP (Tubuline CarboxyPeptidase). Cette enzyme, qui n’avait jusqu’à ce jour jamais été identifiée (sa taille et sa séquence restaient inconnues), a comme fonction de supprimer le dernier acide aminé, une tyrosine, de l’extrémité de la tubuline α. C’est la réaction de détyrosination. Une enzyme réverse, la ligase TTL, est chargée de repositionner cette tyrosine à sa place. C’est la tyrosination. Ce cycle de détyrosination/tyrosination est vital pour la cellule et l’organisme. Une détyrosination massive (anormale) est observée dans plusieurs cancers sévères et maladies cardiaques.
Identifier et caractériser la TCP constituait donc un objectif majeur pour comprendre la fonction physiologique de la détyrosination de la tubuline α et pour évaluer les conséquences de son inhibition.
Pour isoler la TCP, les chercheurs ont suivi son activité, utilisé des techniques classiques de biochimie et fait appel à des chimistes de l’Université de Stanford qui ont développé une petite molécule inhibitrice de son activité. Cette molécule a été utilisée comme hameçon pour « pêcher » l’enzyme convoitée.
Les microtubules sont des fibres présentes dans toutes les cellules composées d’un empilement de tubulines α/β. La tubuline α porte une tyrosine (Y) à son extrémité qui est alternativement enlevée et replacée par deux enzymes, modifiant ainsi la surface des microtubules. La TCP (représentée par une scie composée de deux éléments, VASH/SVBP) est responsable de la détyrosination. La TTL (représentée par un tube de colle) replace la tyrosine sur la tubuline. Ce cycle est essentiel aux diverses fonctions des microtubules dans les cellules (division, migration, …) et vital pour l’organisme. © C. Bosc, GIN
Au final, ce ne sont pas une, mais deux enzymes qui ont été découvertes ! Ces dernières, dénommées VASH1 et VASH2, étaient déjà connues des scientifiques mais sans savoir qu’il s’agissait d’enzymes en lien avec le cytosquelette. Les chercheurs ont montré qu’à la condition d’être associées à une protéine partenaire appelée SVBP, VASH1 et VASH2 sont capables de détyrosiner la tubuline α. Pour le démontrer, les chercheurs ont supprimé leur expression (ou celle de leur partenaire SVBP) dans les neurones. Ils ont alors observé une très forte diminution du taux de détyrosination de la tubuline α, ainsi que des anomalies dans la morphologie des neurones (v. Figure). Les chercheurs sont allés plus loin en montrant que ces enzymes sont également impliquées dans le développement du cortex cérébral.
Des perspectives pour la lutte contre le cancer
Ainsi, quarante ans après les premiers travaux sur la détyrosination de la tubuline α, les enzymes responsables ont été démasquées ! Dorénavant, les scientifiques espèrent qu’en modulant l’efficacité de la TCP et en améliorant les connaissances du cycle détyrosination/tyrosination, ils pourront mieux lutter contre certains cancers et progresseront dans la connaissance des fonctions cérébrales et cardiaques.
[1] Les instituts suivants sont impliqués : Grenoble Institut des neurosciences, GIN (Inserm/Univ. Grenoble Alpes); l’Institut de biosciences et biotechnologies de Grenoble, BIG (Inserm/CEA/Univ. Grenoble Alpes) ; l’Institut pour l’avancée des biosciences, IAB (Inserm/CNRS/Univ. Grenoble Alpes), le Department of Pathology, Stanford University School of Medicine (Stanford, USA), l’Institut de génétique humaine, IGH (CNRS/Univ. de Montpellier), le Centre de recherche en biologie cellulaire de Montpellier, CRBM (CNRS/Univ. de Montpellier).
[2] La tyrosine est l’un des 22 acides aminés qui constituent les protéines
DOCUMENT inserm LIEN |
| |
|
| |
|
| Page : [ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 ] Précédente - Suivante |
|
|
|
|
|
|